

ピックアップ
2026年05月20日【プレスリリース】生化学
東京女子医科大学
大脳皮質樹状突起形成を制御する
新たなリン酸化シグナル機構を解明
東京女子医科大学・生化学分野の若槻実祐(大学院生)、瀧澤光太郎助教、中村史雄教授らの研究グループは、横浜市立大学・先端医科学研究センターの木村鮎子特任助教(現、群馬パース大学)、平野久教授らと共に、大脳皮質樹状突起形成においてチロシンホスファターゼPTPδがSIRPαのリン酸化状態を制御していることを発見しました。脳の神経細胞が複雑なネットワークを形成するためには、樹状突起が正しく伸び、配置される必要があります。しかしその形成を制御する分子メカニズム、特にチロシンリン酸化・脱リン酸化シグナルの役割は十分には解明されていませんでした。
本研究では、チロシンホスファターゼPTPδが、免疫分子として知られるSIRPαのリン酸化状態を精密に制御することで、大脳皮質ニューロンの樹状突起形成を調節していることを明らかにしました。さらに、SIRPαのリン酸化異常をもつマウスでは、神経回路形成に重要な樹状突起の向きや枝分かれに異常が生じることを発見しました。本成果は、脳神経回路がどのように形成されるかという基礎的理解を深めるだけでなく、自閉スペクトラム症や統合失調症など、神経回路形成異常が関与する神経精神疾患の新たな理解につながる可能性があります。
脳の神経細胞は、「樹状突起」と呼ばれる枝状構造を形成することで複雑な神経回路を構築しています。細胞内シグナル伝達では、「チロシンリン酸化」と「脱リン酸化」が重要な役割を果たしています。これらは、それぞれチロシンキナーゼとプロテインチロシンホスファターゼ(PTP)によって制御されます。これまで、チロシンキナーゼ側の研究は数多く進められてきましたが、PTPがどの分子を基質として働き、神経発達にどのように関与しているかについては、十分に明らかになっていませんでした。
PTPδ(PTPRD)は脳に豊富に発現するPTPの一つであり、空間学習やシナプス可塑性に関与することが知られています。また近年では、自閉スペクトラム症、統合失調症、むずむず脚症候群などの神経疾患との関連も報告されています。しかし、PTPδが神経回路形成をどのように制御しているか、その詳細な分子機構は未解明でした。神経回路形成の過程では軸索ガイダンス因子の一つ、Semaphorin 3A(Sema3A)が大脳皮質錐体ニューロンの樹状突起形成を促進することが知られています。PTPδはSema3Aの細胞内シグナル伝達に関与し、Fynチロシンキナーゼを活性化することで、大脳皮質錐体ニューロンの樹状突起形成を促進します(Nakamura F. et al., 2017. J Neurosci 37:7125-7139)。しかし、PTPδがどの分子を脱リン酸化しているのか、すなわち「PTPδの生体内基質」は明らかになっていませんでした。
Signal Regulatory Protein α(SIRPα)は免疫系で“don’t eat me signal”に関与する分子として知られていますが、神経系においても神経細胞の成長円錐やシナプス豊富領域に局在することが報告されています。しかしSIRPαの神経発達における役割はほとんど分かっていませんでした。そこで本研究では、PTPδの新たな基質として同定したSIRPαに着目し、SIRPαのリン酸化・脱リン酸化がSema3A依存性の樹状突起形成にどのように関与するかを解析しました。
PTPδ欠損マウス脳では野生型マウスに比していくつかのタンパク質が過剰にリン酸化されています(図1A)。リン酸化プロテオーム解析により矢印の100kDa蛋白はSignal Regulatory Protein α(SIRPα)であること(図1B)、SIRPαのTyr501残基が著しく過剰リン酸化されていることを見出しました。さらに解析を進めた結果、SIRPαはチロシンキナーゼFynでリン酸化され、PTPδはSIRPαを直接脱リン酸化することから、SIRPαはPTPδの新たな生体内基質であることが示されました。
次に研究グループは、大脳皮質ニューロンの初代培養を用いてSIRPαの役割を解析しました。その結果、PTPδが発現するニューロンではSema3A刺激によってSIRPα Tyr501の脱リン酸化が誘導されることを見出しました(図1C)。またSema3A刺激は野生型ニューロンの樹状突起を伸長させるのに対して、SIRPα欠損ニューロンでは刺激に伴う伸長が消失しました。(図1D左側)。このSIRPα欠損ニューロンに、野生型のSIRPαを再導入すると樹状突起形成は回復しますが(図1D中央)、Tyr501がリン酸化されないSIRPα変異体(Y501F)の導入では回復しませんでした(図1D右側)。生体内解析では、Y501F変異体を発現した大脳皮質錐体ニューロンにおいて、基底樹状突起形成の低下などの異常が認められました(図1E右側)。またSIRPα欠損マウス脳においても皮質ニューロンの樹状突起形成の低下が認められました。
Sema3A刺激による樹状突起伸長の消失や、基底樹状突起の低発達はFynやPTPδの欠損ニューロンでも生じます。これらの結果からFyn, PTPδによるSIRPαのリン酸化・脱リン酸化サイクル、すなわちリン酸化・脱リン酸化の動的変化が樹状突起形成や神経回路発達に重要であることが示されました(図1F)。
次に研究グループは、大脳皮質ニューロンの初代培養を用いてSIRPαの役割を解析しました。その結果、PTPδが発現するニューロンではSema3A刺激によってSIRPα Tyr501の脱リン酸化が誘導されることを見出しました(図1C)。またSema3A刺激は野生型ニューロンの樹状突起を伸長させるのに対して、SIRPα欠損ニューロンでは刺激に伴う伸長が消失しました。(図1D左側)。このSIRPα欠損ニューロンに、野生型のSIRPαを再導入すると樹状突起形成は回復しますが(図1D中央)、Tyr501がリン酸化されないSIRPα変異体(Y501F)の導入では回復しませんでした(図1D右側)。生体内解析では、Y501F変異体を発現した大脳皮質錐体ニューロンにおいて、基底樹状突起形成の低下などの異常が認められました(図1E右側)。またSIRPα欠損マウス脳においても皮質ニューロンの樹状突起形成の低下が認められました。
Sema3A刺激による樹状突起伸長の消失や、基底樹状突起の低発達はFynやPTPδの欠損ニューロンでも生じます。これらの結果からFyn, PTPδによるSIRPαのリン酸化・脱リン酸化サイクル、すなわちリン酸化・脱リン酸化の動的変化が樹状突起形成や神経回路発達に重要であることが示されました(図1F)。
Ⅲ 今後の展開
本研究により、PTPδによるSIRPαのリン酸化制御が、大脳皮質ニューロンの樹状突起形成に重要であることが明らかとなりました。今後は、SIRPαのリン酸化・脱リン酸化が樹状突起形成においてどのように時空間的に制御されているかを解明し、さらにPTPδの活性制御機構の解明を目指します。また、PTPRD遺伝子異常は自閉スペクトラム症や統合失調症などとの関連が報告されていることから、本研究成果は、これら神経発達疾患の新たな分子病態理解や治療戦略の開発につながることが期待されます。
本研究により、PTPδによるSIRPαのリン酸化制御が、大脳皮質ニューロンの樹状突起形成に重要であることが明らかとなりました。今後は、SIRPαのリン酸化・脱リン酸化が樹状突起形成においてどのように時空間的に制御されているかを解明し、さらにPTPδの活性制御機構の解明を目指します。また、PTPRD遺伝子異常は自閉スペクトラム症や統合失調症などとの関連が報告されていることから、本研究成果は、これら神経発達疾患の新たな分子病態理解や治療戦略の開発につながることが期待されます。
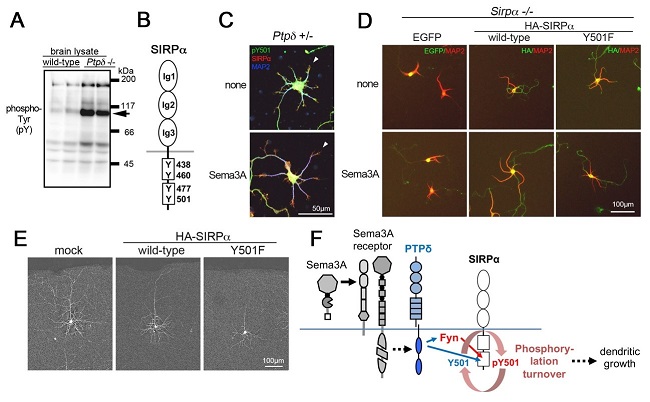
図1. PTPδによるSIRPα脱リン酸化はSema3A誘導性樹状突起形成を制御する。
A) 野生型(wild-type)、PTPδ欠損(Ptpδ -/-)脳におけるリン酸化チロシンブロット。PTPδ欠損脳において複数タンパク質が過剰にリン酸化されている。100kDaタンパク質はプロテオーム解析によりSIRPαと同定された。B) SIRPα模式図。細胞外に3つのイムノグロブリン様領域、細胞内に4つのチロシン残基(Y)を有する。数字はアミノ酸の位置。C) PTPδを発現するニューロンの初代培養。樹状突起(矢頭)の先端にリン酸化SIRPαの緑シグナル(pY501)とSIRPαの赤シグナルが共在し黄色になる。Sema3A刺激により樹状突起のSIRPαは脱リン酸化され、赤シグナルのみになる(矢頭)。D) SIRPα欠損(Sirpα -/-)ニューロンの初代培養。Sema3Aによる樹状突起(赤の突起)伸長は生じない(左列)。SIRPα野生型を再発現させると、Sema3A刺激により樹状突起が伸長する(中央列)。SIRPαリン酸化変異体(Y501F)の発現では、Sema3Aによる樹状突起伸長は生じない。E) SIRPα野生型、Y501F変異型を導入した大脳皮質ニューロンの樹状突起。SIRPα野生型(中央)の導入は空ベクター(mock, 左側)導入と同様に樹状突起が発達する。一方、Y501F変異体導入は樹状突起発達を抑制した(右側)。F) SIRPαのリン酸化・脱リン酸化による樹状突起伸長模式図。Sema3AはSema3A受容体に結合し、PTPδを活性化する。PTPδはFynを活性化し、FynはSIRPαをリン酸化する。またPTPδはSIRPαを脱リン酸化することから、リン酸化・脱リン酸化の代謝回転の増加が樹状突起の伸長を引き起こすと考えられる。
A) 野生型(wild-type)、PTPδ欠損(Ptpδ -/-)脳におけるリン酸化チロシンブロット。PTPδ欠損脳において複数タンパク質が過剰にリン酸化されている。100kDaタンパク質はプロテオーム解析によりSIRPαと同定された。B) SIRPα模式図。細胞外に3つのイムノグロブリン様領域、細胞内に4つのチロシン残基(Y)を有する。数字はアミノ酸の位置。C) PTPδを発現するニューロンの初代培養。樹状突起(矢頭)の先端にリン酸化SIRPαの緑シグナル(pY501)とSIRPαの赤シグナルが共在し黄色になる。Sema3A刺激により樹状突起のSIRPαは脱リン酸化され、赤シグナルのみになる(矢頭)。D) SIRPα欠損(Sirpα -/-)ニューロンの初代培養。Sema3Aによる樹状突起(赤の突起)伸長は生じない(左列)。SIRPα野生型を再発現させると、Sema3A刺激により樹状突起が伸長する(中央列)。SIRPαリン酸化変異体(Y501F)の発現では、Sema3Aによる樹状突起伸長は生じない。E) SIRPα野生型、Y501F変異型を導入した大脳皮質ニューロンの樹状突起。SIRPα野生型(中央)の導入は空ベクター(mock, 左側)導入と同様に樹状突起が発達する。一方、Y501F変異体導入は樹状突起発達を抑制した(右側)。F) SIRPαのリン酸化・脱リン酸化による樹状突起伸長模式図。Sema3AはSema3A受容体に結合し、PTPδを活性化する。PTPδはFynを活性化し、FynはSIRPαをリン酸化する。またPTPδはSIRPαを脱リン酸化することから、リン酸化・脱リン酸化の代謝回転の増加が樹状突起の伸長を引き起こすと考えられる。
【お問い合わせ先】
<研究に関すること>
中村 史雄(ナカムラ フミオ)
東京女子医科大学 医学部生化学分野 教授
〒162-8666 東京都新宿区河田町8-1
Tel&Fax:03-3353-8112 (Ext: 31525) 直通 03-5269-7415
E-mail: nakamura.fumio@twmu.ac.jp
<報道に関すること>
東京女子医科大学 広報課
〒162-8666 東京都新宿区河田町8-1
Tel:03-3353-8111 Fax:03-3353-6793
E-mail: kouhou.bm@twmu.ac.jp
【プレス情報】
1.掲載誌名:Journal of Neuroscience
2.論文タイトル: Elaboration of cortical pyramidal dendrites requires dephosphorylation of Signal Regulatory Protein α by Protein Tyrosine Phosphatase δ
3.著者名: Miyu Wakatsuki, Kohtaro Takizawa, Aoi Jitsuki-Takahashi, Ayuko Kimura, Nobuto Arashiki, Hiroko Makihara, Yoichiro Iwakura, Takuya Takahashi, Hisashi Hirano, Yoshio Goshima, Fumio Nakamura*
(*はcorresponding author、アンダーラインは本学所属の著者)
4.DOIコード:10.1523/JNEUROSCI.2043-25.2026
5.論文のオンライン掲載日と報道解禁日(Embargo) :2026年5月20日
中村 史雄(ナカムラ フミオ)
東京女子医科大学 医学部生化学分野 教授
〒162-8666 東京都新宿区河田町8-1
Tel&Fax:03-3353-8112 (Ext: 31525) 直通 03-5269-7415
E-mail: nakamura.fumio@twmu.ac.jp
<報道に関すること>
東京女子医科大学 広報課
〒162-8666 東京都新宿区河田町8-1
Tel:03-3353-8111 Fax:03-3353-6793
E-mail: kouhou.bm@twmu.ac.jp
【プレス情報】
1.掲載誌名:Journal of Neuroscience
2.論文タイトル: Elaboration of cortical pyramidal dendrites requires dephosphorylation of Signal Regulatory Protein α by Protein Tyrosine Phosphatase δ
3.著者名: Miyu Wakatsuki, Kohtaro Takizawa, Aoi Jitsuki-Takahashi, Ayuko Kimura, Nobuto Arashiki, Hiroko Makihara, Yoichiro Iwakura, Takuya Takahashi, Hisashi Hirano, Yoshio Goshima, Fumio Nakamura*
(*はcorresponding author、アンダーラインは本学所属の著者)
4.DOIコード:10.1523/JNEUROSCI.2043-25.2026
5.論文のオンライン掲載日と報道解禁日(Embargo) :2026年5月20日
