

お知らせ
2026年03月02日 【プレスリリース】網膜の血管地図を再現する数理モデル-目の病気の理解を前へ-
九州大学
東京女子医科大学
東京女子医科大学
網膜の血管地図を再現する数理モデル
-目の病気の理解を前へ-
| Point
① ヒト網膜には、中心窩*1を取り囲む中心窩無血管域(FAZ*2)など、臨床的に重要な特徴構造があるが、霊長類での実験的制約から形成メカニズムの理解が遅れている。
② 血管新生(VEGF*3勾配に従う頂端細胞*4運動)と、血管の足場となる網膜アストロサイト分布*5の時間発展を結合したモデルにより、ヒト網膜に特徴的な複数の血管パターンを同時に再現した。 ③ 本モデルは、未熟児網膜症(ROP)など網膜疾患に見られる異常血管パターン形成の理解・仮説検証にも応用できる可能性がある。 |
【概要】
ヒト網膜には黄斑*6・中心窩があり、その中心には血管が存在しない領域である中心窩無血管域(FAZ)が形成されます。FAZの形状や周囲血管の配列は視機能や疾患とも関わりますが、霊長類網膜の実験的アプローチが難しいため、ヒト特有の血管構造がどのように形成されるかは十分に解明されていません。
九州大学と東京女子医科大学の共同研究グループは、血管新生と網膜アストロサイトの分布拡張を結合した数理モデルにより、FAZを含むヒト網膜に特徴的な血管パターン形成過程を説明できることを示しました。
九州大学大学院医学系学府系統解剖学分野の吉村公太朗大学院生(研究当時)と同大学院医学研究院系統解剖学分野の三浦岳教授、東京女子医科大学眼科の飯田知弘教授と丸子一朗准教授の共同研究にて、光干渉断層血管撮影(OCTA*7)画像解析と数理モデル構築を行いました。研究グループは、血管先端の細胞がランダムウォークと化学走化性により移動し、その軌跡に血管が形成されるという仮定に基づき、本モデルを構築しました。さらに、血管形成の足場となる網膜アストロサイトが時間とともに拡張し、中心窩近傍では阻害因子の存在により網膜アストロサイトの分布が制限される、という仮定を導入しました。
この結合モデルは、視神経円板(OD*8)から放射状に拡がる血管、上下の耳側アーケード血管*9、FAZ、FAZへ向かう求心的走行、FAZ耳側で上下からかみ合うように並ぶ血管構造など、ヒト網膜で観察される複数の特徴を同時に再現しました。特に、アストロサイト分布の“拡張のタイミング”がFAZ周囲パターン形成に重要であることが示唆され、疾患(未熟児網膜症など)における異常パターン形成の理解・検証に利用できる可能性があります。
本研究成果は、米国の科学雑誌『Translational Vision Science & Technology』(TVST)に2026年3月2日(月)午後10時30分(日本時間)に掲載されます。
本研究グループからひとこと:
実験で追うことが難しいヒト網膜の発生過程を、数理モデルで“ビデオを再生する”ように検討できる枠組みを目指しました。アストロサイトの拡がり方が血管の“道筋”を規定するという見方は、疾患の異常パターン理解にもつながると考えます。
網膜血管発生はこれまで主にマウスを用いて研究され、網膜アストロサイトがODから網膜内へ先行して拡がり、その後に内皮細胞がVEGF勾配に沿って進展して血管網を形成する、という枠組みが明らかにされてきました。一方で、ヒト網膜には黄斑と中心窩が存在し、その中心には血管が存在しない中心窩無血管域(FAZ)が形成されるなど、マウスには見られない特徴的な構造があります。FAZの形状やその周囲の血管配列は視機能や病態とも深く関わると考えられますが、霊長類網膜の発生過程を実験的に追跡することには制約が大きく、ヒト特有の血管パターンがどのように生じるのかは十分に解明されていません。
近年、光干渉断層血管撮影(OCTA)の発展により、従来法では捉えにくかった毛細血管を含む網膜血管構造を非侵襲的に高解像度で観察できるようになりました。これにより、観察されたパターンを定量化し、それと整合する形成メカニズムを数理モデル側から検証することが現実的になってきました。しかし、これまでに提案されてきた血管新生の数理モデルは腫瘍血管などを対象とした枠組みが中心であり、ヒト網膜発生、とりわけFAZ周囲の特徴的構造を説明するモデルは確立していませんでした。そこで本研究では、ヒト網膜血管の形成過程、特にFAZ周囲のパターンがどのように立ち上がるのかを、OCTAによる観察像と照合しながら数理的に検討することを目的としました。
近年、光干渉断層血管撮影(OCTA)の発展により、従来法では捉えにくかった毛細血管を含む網膜血管構造を非侵襲的に高解像度で観察できるようになりました。これにより、観察されたパターンを定量化し、それと整合する形成メカニズムを数理モデル側から検証することが現実的になってきました。しかし、これまでに提案されてきた血管新生の数理モデルは腫瘍血管などを対象とした枠組みが中心であり、ヒト網膜発生、とりわけFAZ周囲の特徴的構造を説明するモデルは確立していませんでした。そこで本研究では、ヒト網膜血管の形成過程、特にFAZ周囲のパターンがどのように立ち上がるのかを、OCTAによる観察像と照合しながら数理的に検討することを目的としました。
【研究の内容と成果】
本研究ではまず、FAZ形成に関与し得る要因として想定される四つの仮説を検証しました。すなわち、中心窩から内皮阻害因子が分泌される仮説、中心窩で内皮誘引因子が欠損する仮説、組織変形に伴って血管が中心窩から引き離される仮説、そして中心窩への進入が単純に遮断される仮説です。これらを、頂端細胞の運動に基づく血管新生モデルの枠組みの中で実装し比較したところ、FAZを避ける現象そのものは生じても、ヒト網膜に特徴的な「FAZへ向かう求心的走行」や「FAZ耳側で上下からかみ合うように配列する構造」などを十分に再現できませんでした。
次に、血管新生の過程に網膜アストロサイトの効果を明示的に組み込みました。数理モデルでは、血管先端である内皮頂端細胞がランダムウォークと化学走化性により移動し、その軌跡に血管が形成されると仮定しました。化学走化性因子としてはVEGFを想定し、低酸素ほどVEGF発現が高いという関係を踏まえて、計算では酸素勾配を代替指標として用いました。さらに、網膜アストロサイトが時間とともに網膜内へ拡張し、その分布域が血管形成の足場となって、頂端細胞は網膜アストロサイトが存在する領域でのみ進める、という仮定を導入しました。加えて、発生中の中心窩領域に網膜アストロサイトが分布しにくいという観察事実を反映させるため、中心窩近傍に網膜アストロサイト拡張を抑制する阻害因子の分布を仮定しました。
このように血管新生と網膜アストロサイト動態を結合させた数理モデルは、ODから放射状に拡がる血管構造、上下の耳側アーケード血管、FAZ構造、FAZへ向かう求心的走行、そしてFAZ耳側で上下から対面し境界が形成されるような配列など、ヒト網膜に特徴的な複数の構造を同時に再現しました。特に、網膜アストロサイト分布を時間的に固定してしまうとFAZ周囲の特徴が弱まることから、網膜アストロサイトの「存在」だけではなく「拡張のタイミング」を含む動態そのものがパターン形成に本質的であることが示唆されました。
【今後の展開】
本研究で示した枠組みは、生体内で直接観察することが難しいヒト網膜血管の発生過程を、数理モデル上で時間発展として提示できる点に強みがあります。したがって、未熟児網膜症をはじめとする疾患で見られる異常血管パターンについて、酸素環境や分枝条件、網膜アストロサイト動態の遅れや変調といった仮説を、計算機上で系統的に検証するための基盤として応用できる可能性があります。将来的には、血流に伴うリモデリングや血管径の階層性、網膜外からの酸素供給など、現実の要素をモデルへ段階的に組み込み、再現性と予測力を高めることが重要です。また、OCTA画像から推定される特徴量とモデルパラメータを接続することで、画像に基づく「解釈可能な」メカニズム推定へ展開できれば、発生異常や疾患病態の理解に資する枠組みへ発展し得ると考えられます。
【参考図】
本研究ではまず、FAZ形成に関与し得る要因として想定される四つの仮説を検証しました。すなわち、中心窩から内皮阻害因子が分泌される仮説、中心窩で内皮誘引因子が欠損する仮説、組織変形に伴って血管が中心窩から引き離される仮説、そして中心窩への進入が単純に遮断される仮説です。これらを、頂端細胞の運動に基づく血管新生モデルの枠組みの中で実装し比較したところ、FAZを避ける現象そのものは生じても、ヒト網膜に特徴的な「FAZへ向かう求心的走行」や「FAZ耳側で上下からかみ合うように配列する構造」などを十分に再現できませんでした。
次に、血管新生の過程に網膜アストロサイトの効果を明示的に組み込みました。数理モデルでは、血管先端である内皮頂端細胞がランダムウォークと化学走化性により移動し、その軌跡に血管が形成されると仮定しました。化学走化性因子としてはVEGFを想定し、低酸素ほどVEGF発現が高いという関係を踏まえて、計算では酸素勾配を代替指標として用いました。さらに、網膜アストロサイトが時間とともに網膜内へ拡張し、その分布域が血管形成の足場となって、頂端細胞は網膜アストロサイトが存在する領域でのみ進める、という仮定を導入しました。加えて、発生中の中心窩領域に網膜アストロサイトが分布しにくいという観察事実を反映させるため、中心窩近傍に網膜アストロサイト拡張を抑制する阻害因子の分布を仮定しました。
このように血管新生と網膜アストロサイト動態を結合させた数理モデルは、ODから放射状に拡がる血管構造、上下の耳側アーケード血管、FAZ構造、FAZへ向かう求心的走行、そしてFAZ耳側で上下から対面し境界が形成されるような配列など、ヒト網膜に特徴的な複数の構造を同時に再現しました。特に、網膜アストロサイト分布を時間的に固定してしまうとFAZ周囲の特徴が弱まることから、網膜アストロサイトの「存在」だけではなく「拡張のタイミング」を含む動態そのものがパターン形成に本質的であることが示唆されました。
【今後の展開】
本研究で示した枠組みは、生体内で直接観察することが難しいヒト網膜血管の発生過程を、数理モデル上で時間発展として提示できる点に強みがあります。したがって、未熟児網膜症をはじめとする疾患で見られる異常血管パターンについて、酸素環境や分枝条件、網膜アストロサイト動態の遅れや変調といった仮説を、計算機上で系統的に検証するための基盤として応用できる可能性があります。将来的には、血流に伴うリモデリングや血管径の階層性、網膜外からの酸素供給など、現実の要素をモデルへ段階的に組み込み、再現性と予測力を高めることが重要です。また、OCTA画像から推定される特徴量とモデルパラメータを接続することで、画像に基づく「解釈可能な」メカニズム推定へ展開できれば、発生異常や疾患病態の理解に資する枠組みへ発展し得ると考えられます。
【参考図】
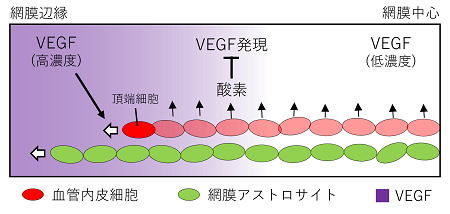
図1 数理モデル模式図
血管伸長に先行して網膜アストロサイト(緑楕円)が拡がり、これを足場として頂端細胞(Tip cell)が運動した経路上に血管構造(赤楕円)が形成されると仮定した。この頂端細胞の運動は、血管から供給される酸素によって決定されるVEGF分布の勾配に誘引されると仮定した。
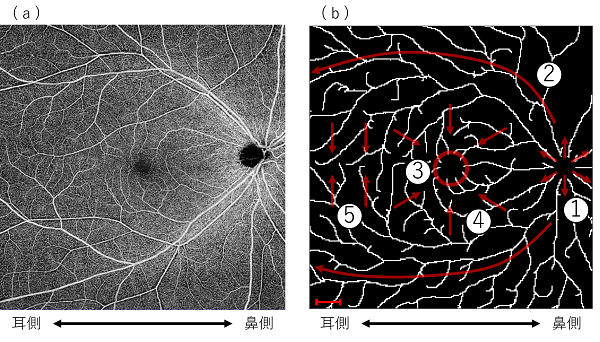
図2 OCTA画像と数理モデル結果図の比較
血管伸長に先行して網膜アストロサイト(緑楕円)が拡がり、これを足場として頂端細胞(Tip cell)が運動した経路上に血管構造(赤楕円)が形成されると仮定した。この頂端細胞の運動は、血管から供給される酸素によって決定されるVEGF分布の勾配に誘引されると仮定した。
図2 OCTA画像と数理モデル結果図の比較
(a): OCTA画像。(b): 数理モデル結果図。研究グループが構築した数理モデルは、① ODから放射状に拡がる血管構造、② 上下の耳側アーケード血管、③FAZ構造、④ FAZへ向かう求心的走行、⑤ FAZ耳側で上下から対面し境界が形成されるような配列といった複数の構造的特徴を再現した。
【用語解説】
(*1) 中心窩(ちゅうしんか):視野の中心に該当する網膜領域に存在する、網膜の層構造が薄くなっている領域。
(*2) 中心窩無血管域(FAZ):中心窩の周囲に形成される、血管が存在しない領域。
(*3) 血管内皮増殖因子(VEGF):低酸素環境で発現が増加し、血管新生を促進する因子。
(*4) 頂端細胞(Tip cell):血管新生の先端で伸長を先導する内皮細胞。周囲の環境(化学走化性など)に応答して移動し、血管の伸長方向を決める。
(*5) 網膜アストロサイト:神経系のグリア細胞。発生期網膜では血管伸長の足場提供や因子分泌などを通して血管形成を支える役割が示唆されている。
(*6) 黄斑:明所での視力や色覚を担う錐体細胞が高密度に存在する網膜の中心領域。
(*7) 光干渉断層血管撮影(OCTA):光干渉断層計(OCT)を応用し網膜血管など眼底の血管に特化した撮像法。OCT信号の時間変化から血流を推定し、網膜血管を非侵襲的に可視化する。
(*8) 視神経円板(OD):視神経乳頭(ONH)とも呼ばれる、視神経繊維が網膜から脳へつながる際の“出口”となっている網膜上の構造。網膜内の動静脈の“出入口”でもあり、ヒトではやや鼻側に存在する。
(*9) 耳側アーケード血管:黄斑の上下を取り囲むようにODから弓なりに伸びる血管構造。
【謝辞】
本研究では、植村明嘉氏(名古屋市立大学)に有益な助言や議論をしていただきました。この場を借りて深く御礼申し上げます。
【論文情報】
掲載誌:Translational Vision Science & Technology
タイトル:Mathematical modeling of human retinal vascular pattern around the foveal avascular zone
著者名:Kotaro Yoshimura, Kei Sugihara, Ichiro Maruko, Tomohiro Iida, Takashi Miura
DOI:10.1167/tvst.15.3.1
(*1) 中心窩(ちゅうしんか):視野の中心に該当する網膜領域に存在する、網膜の層構造が薄くなっている領域。
(*2) 中心窩無血管域(FAZ):中心窩の周囲に形成される、血管が存在しない領域。
(*3) 血管内皮増殖因子(VEGF):低酸素環境で発現が増加し、血管新生を促進する因子。
(*4) 頂端細胞(Tip cell):血管新生の先端で伸長を先導する内皮細胞。周囲の環境(化学走化性など)に応答して移動し、血管の伸長方向を決める。
(*5) 網膜アストロサイト:神経系のグリア細胞。発生期網膜では血管伸長の足場提供や因子分泌などを通して血管形成を支える役割が示唆されている。
(*6) 黄斑:明所での視力や色覚を担う錐体細胞が高密度に存在する網膜の中心領域。
(*7) 光干渉断層血管撮影(OCTA):光干渉断層計(OCT)を応用し網膜血管など眼底の血管に特化した撮像法。OCT信号の時間変化から血流を推定し、網膜血管を非侵襲的に可視化する。
(*8) 視神経円板(OD):視神経乳頭(ONH)とも呼ばれる、視神経繊維が網膜から脳へつながる際の“出口”となっている網膜上の構造。網膜内の動静脈の“出入口”でもあり、ヒトではやや鼻側に存在する。
(*9) 耳側アーケード血管:黄斑の上下を取り囲むようにODから弓なりに伸びる血管構造。
【謝辞】
本研究では、植村明嘉氏(名古屋市立大学)に有益な助言や議論をしていただきました。この場を借りて深く御礼申し上げます。
【論文情報】
掲載誌:Translational Vision Science & Technology
タイトル:Mathematical modeling of human retinal vascular pattern around the foveal avascular zone
著者名:Kotaro Yoshimura, Kei Sugihara, Ichiro Maruko, Tomohiro Iida, Takashi Miura
DOI:10.1167/tvst.15.3.1
【お問い合わせ先】
<研究に関すること>
九州大学 大学院医学研究院 系統解剖学分野 教授 三浦 岳(ミウラ タカシ)
TEL:092-642-6048 FAX:092-642-6923
Mail:miura.takashi.869@m.kyushu-u.ac.jp
東京女子医科大学 医学部眼科学講座 准教授 丸子 一朗(マルコ イチロウ)
TEL/FAX:03-3353-8111
Mail:imaruko@twmu.ac.jp
<報道に関すること>
九州大学 広報課
TEL:092-802-2130 FAX:092-802-2139
Mail:koho@jimu.kyushu-u.ac.jp
東京女子医科大学 広報課
TEL:03-3353-8111 FAX:03-3353-6793
Mail:kouhou.bm@twmu.ac.jp
